水下仿生机器人尾鳍振荡产生的推力建模∗
尹欣妍,贾立超™,王晨,谢光明
湍流与复杂系统国家重点实验室
北京大学工程系,北京100871
摘要:基于水下仿生机器人尾鳍的振动,提出了推力的简化模型。用中心模式发生器(CPGs)对尾鳍振荡进行了推广。在该模型中,阻力系数和升力系数是通过数字粒子图像测速(DPIV)和力传感器实验得到的两个关键参数。通过数值模拟和物理实验对该动力学模型进行了验证。
关键词尾鳍振荡,推力,水下仿生机器人,数字粒子图像测速(DPIV),中心模式发生器(CPG)
1 介绍
人们常认为鱼是游泳能手。了解和适应鱼类游泳运动的基本原理,使之适应人工游泳机器,对学术界和工业界都有很大的吸引力。通常,鱼类利用身体/尾鳍(BCF)波动来推进[1]。不同类型的BCF物种在运动和水动力性能上表现出明显的多样性。建立合适的动力学模型是优化仿生设计的关键。
据我们所知,鱼类的游泳能力在许多方面都远远超过了数百万年来航海科学和技术在广阔且常常充满敌意的领域所取得的成就[2]。采用静态流体理论,采用准静态近似方法建立了游泳运动的生物阻力理论(泰勒),对鱼类的游泳机理进行了研究。Lighthill[4]提出了一个拉长的carangidae模型,强调小体积水与动物表面接触部分之间的反作用力。吴〔5〕提出了二维柔性板理论,将鱼作为二维弹性薄板。Weihs将大振幅细长体理论(见文献[6])和机翼升力法相结合,计算了敏捷游泳时的力,并认为该力相当于尾迹涡的力[7-8]。最近,Yang等人[9]为研究自由自航鱼类提供了变形体动力学和非定常流体动力学的综合方法。
许多研究人员利用数字粒子图像测速技术(DPIV)对鱼类尾迹流场进行了观测和研究。Lauder和Drucker[10]使用DPIV观测尾迹流场,并解释了每个鳍的功能。Anderson等人[11]通过研究流场的可视化,发现攻角是影响推进效率的一个重要参数。Wen等人[12]使用DPIV测量来说明机器人游泳者(Angulliform、carangiform和thunniform)产生的流型特征,此外,提出了一种力反馈控制的实验方法。
我们将生物行为与水下仿生机器人进行了整合,并进一步评估了可能的结果,以实现可靠的控制。有一些研究集中在面向控制的游泳建模上。Ekeberg[13]提出了一个模拟水中作用力的力学模型。该模型基于两个假设。一个是雷诺数足够高,惯性力占主导地位,另一个是流体是静止的。McIsaac和Ostrowski在文献[14]中提出了对称结构类鳗鱼机器人的Lagrangian模型。Boyer等人[15]提出了一个连续的三维(3D)游动类鳗鱼机器人的动力学模型。Ding等人[16]利用拉格朗日方法提出了一个动力学模型,并实现了向后游动。然而,上述模型的形式都比较复杂,限制了它们在工程中的应用。
本文建立了基于中心模式发生器(CPGs)控制的多关节水下机器人动力学模型。此外,还进行了实验研究,建立了该水下机器人尾鳍振荡与推力之间的简单数学模型。并与数值模拟结果进行了比较,验证了模型的正确性,结果表明两者之间存在可接受的差异。
论文的其余部分安排如下。第二节介绍了该模型和基于CPG的机车控制。第三节介绍了水下机器人的测量方法和动力学模型的建立。第四节给出了仿真和实验结果。最后,在最后一部分给出了本文的结论和未来工作的展望。
2 基于CPG的机车控制模型
考虑到自由游动动物实验研究的难度,采用多连杆仿生机器人实现鱼样波动,如图1(a)所示。该机器人被建模为一个由四个相同的链节组成的链,链节由三个旋转关节串联而成。为了估计鱼体的振荡特性,Lighthill的细长体理论被广泛应用于产生推进行波。同时,许多其他方面,如简单的时间指数正弦函数被用来产生行波。此外,由于系统抗干扰能力差、控制参数突变等缺点,会导致推进波的不连续,进而产生非光滑的轨迹。因此,CPG是机器人的基本控制模型,它可以保证游泳时的基本节奏运动。
cpg是一种神经网络,它能够产生协调的节律活动模式,而无需任何来自感觉反馈或更高控制中心的节律输入[17–18],近年来已广泛应用于机器人领域。控制器系统由三个耦合线性振荡器组成,第i(i=1,2,3)个振荡器的实现如下:
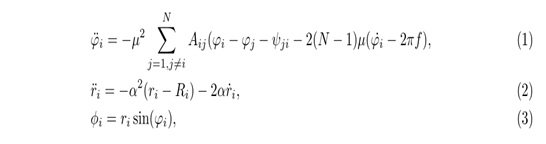
如表1所示,参数是用CPG方法生成的。所有参数均应用于实验和数值模拟中。图1(b)描绘了三个关节的节律性CPG输出信号。
3.1 测量技术
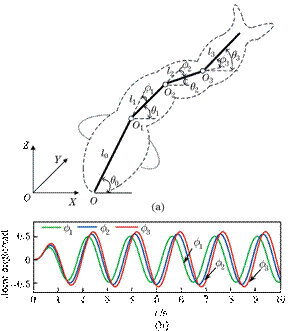
为了确定模型中使用的参数,采用了两种不同的测量方法,即DPIV[19–22]和力传感器,观察流场并计算推力Ft。这两种测量均在静水池中进行,假设推力主要来自尾鳍与周围环境的相对运动首先,利用DPIV测量尾翼产生的尾迹流场,估计尾翼的平均推力。正如生物学家所建议的,尾鳍中线是进行二维尾流测量的最佳水平位置[23]。假设三维涡环的深度等于尾鳍的高度,而其他水平面上的测量平面低估了推力[24]。图2给出了实验装置的概貌。机器鱼可以通过射频模块由计算机控制。DPIV系统的激光板平面被设置为穿过机器人尾鳍的中心线。中径为10μm的粒子被激光片照射。互补金属氧化物半导体(CMOS)相机(1024×1024像素)捕获了拍摄频率为50Hz的流场。采用戴尔计算机控制整个DPIV系统,同时存储粒子图像。波长为532nm的双脉冲激光器(15mJ/脉冲)被圆柱形透镜扩展以产生光片。长焦距镜头用于调节灯片腰部厚度约1mm。由于尾鳍的位置包含在粒子图像中,因此鱼群的游动行为也可以由DPIV结果进行分析。
由于DPIV的时间分辨率限制在50Hz,单分量力传感器HBM Z6FC3用于测量静水中游泳机器人的推力,其采样率高达5 kHz,灵敏度在轴向方向为0.1Mn。测量前进行静态校准。力标定结果表明,前向力的线性度优于1%。然后,将力传感器垂直装配在机器人上方,以测量机器鱼的轴向力。外力传感器的输出由计算机通过国家仪器(NI)前置放大器记录。同步信号用于触发力传感器和摄像机的数据采集。然后,尾鳍后面的流动结构图像,与力的测量和鱼的运动同步。
3.2 动力分析
当机器人在水中游泳时,作用在身体上的力取决于身体和周围环境之间的相对运动。在相对较低的速度下,粘性力在决定运动状态方面起着重要的作用。同时,对于更高速度的情况,惯性效应成为主导因素[25]。根据实验结果,雷诺数(Rel=U l/ν)为105,其大小足以忽略粘性力的影响,其中l为体长,U和ν分别为机器人的自推进速度和水的运动粘度。
本着简化的精神,在不考虑侧向力的情况下,将作用在机器人上的力分为阻力和推力。阻力D通常相当于具有相同雷诺数Re和流速的刚体的流体阻力。此外,拖曳力D的方向是沿着身体轴线从头部到尾部,
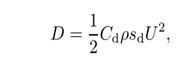
式中,ρ为流体密度,sd为机器人本体的有效截面积,Cd为随Re变化的阻力系数,实际通过实验确定。这里,Cd被选为[26]
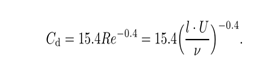
根据细长体理论,推力主要由尾鳍产生[27]。这使得推力Ft被估计为施加在尾鳍上的反作用力是合理的。如Li和Yin[28]所示,不同行为的尾鳍模型的力具有相似的形状。然后,推力可以表示为:
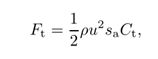
其中u是尾翼的特征速度,sa是尾翼的面积,Ct是推力系数[25]。
如之前的研究所示,无量纲参数Strouhal数支配鱼类运动的水动力性能[22–23,28],定义如下:
其中A表示尾鳍尖的波动幅度。根据式(6),推力系数Ct可表示如下:
.
一个显著的事实是,这里使用了特征速度u的非传统定义,即尾翼的平均摆动速度。这种速度在计算推力时更为合理,本文后面将对此进行解释。特征速度u是指鳍在一个圆内的平均速度,即。,
然后,很容易得出特征速度u是游泳速度和Strouhal数的某种组合,即。,
4 结果和讨论
图3显示了机器人尾鳍在不同拍打频率下产生的流场的DPIV时间序列。在一个拍打周期内,尾鳍在每次拍打时产生一个涡环。图3所示的尾迹结构可归因于双排涡,Strouhal数分别为0.77和0.59。这与之前的实验研究[29-30]一致,其结果表明,当0.325<St<1.025时,机器鱼产生的尾迹是“双排街”。如图3(a)所示,涡是从左侧(朝右侧)产生的,形成涡环R1,用虚线包络线表示。尾鳍向右移动时,涡强度逐渐增强,这意味着机器鱼在椭圆涡环中的冲量在增加。从图3(b)可以看出,涡1和涡2是由右旋(向左)产生并形成涡环R1,涡3和涡4是由左旋产生并形成涡环R2。与图3(a)相比,图3(b)的涡流强度越来越强,因为剪切强度会随着弹跳频率的增加而增加。
图3靠近左侧的短线表示机器人尾鳍的位置,白色箭头表示拍打方向,涡流数位于涡流中心附近:(a)双排涡流,记录在St=0.77,f=0.9Hz,Δt=0.1T;(b)双排涡流,记录在St=0.59,f=1.5Hz,Δt=0.5T
涡流动力学模型假设机器人释放的所有脉冲都包含在椭圆涡流环中,可以用来分析尾迹,以获得机器人的推力[12]。流场是通过冻结每个漩涡在其梭口位置形成的。然后,利用二维涡动力学理论计算了测量尾迹的冲量,用于推力的确定。
在实验过程中,尾鳍的振幅在不同频率下并不总是恒定的。对于较低的拍打频率,尾鳍的设计振幅可以达到。当频率超过阈值时,尾鳍的振幅随振荡频率的增加而减小。这可以通过转向器响应的限制和尾鳍水动力负荷的增加来解释。用尾鳍振幅A的平方和振荡频率的乘积来描述这两个变量之间的关系。如图4所示,当拍打频率大于
本实验为0.7Hz。
图4不同情况下尾鳍振幅和振荡频率平方和f的乘积
这一趋势可以在三个不同的控制参数下观察到。这意味着,如果阈值频率可以确定的话,水下仿生旋翼的实际运动参数将根据不同的控制情况由该定义的产品导出。
以案例3为例。瞬时无量纲推力系数Ct结果如式(8)所示,如图5(a)所示。不同频率下的无量纲推力系数曲线,在形状和平均值基本相同的情况下,会发生塌陷。类似的趋势可以从先前的结果中得到[12]。在我们的实验中,不同频率的平均无量纲推力系数在0.64±0.06范围内,如图5(b)所示。在相同的拍打频率下,特征速度大于游动速度。因此,不同参考速度的标准化无量纲推力系数之间的比较如图5(c)所示。图8(b)显示了不同频率下相应的实验游泳速度。结果表明,与机器人的自推进速度相比,本文采用的特征速度更适合于推力的计算。这一事实从另一个角度证明了Strouhal数是鱼类运动水动力性能的主导无量纲参数。
RASPA等人〔29〕通过对起伏薄的柔性箔片的游泳动力学进行实验研究来解决阻力的建立。结果表明,在机翼侧缘卷起的尾涡对总阻力的产生起着重要作用。对于所有展弦比,单位跨距的箔片运动推力相同。这里使用的长宽比(R)是游泳者尾巴的跨度和长度之间的比值。然而,在计算图6(a)所示的林分无量纲净推力系数Ct时,存在一些差异。如果我们用尾鳍的平均摆动速度而不是身体的自推进速度作为特征速度,Ct值就会变得更加集中,如图6(b)所示。
为了深入了解这种关系,我们估计了机器人的向前游动。类鱼游泳的固有波动特性提供瞬时推力(Ft)、阻力(Fd)和净力(Fnet),如图7所示。仿真和真实机器人自由泳速度的比较如图8所示。
根据实验结果,我们假设Ct可以表示为
C类t型=C0+Ca cos(2πf),(11)
式中常系数C0,Ca用最小二乘法优化。
图5(a)瞬时无量纲推力系数Ct;(b)作为频率函数的平均无量纲推力系数Ct;(c)不同参考速度的标准化无量纲推力系数Ct
图6(a)推进速度为特征速度[27],(b)尾鳍平均摆动速度为特征速度的无量纲推力系数
结果表明,虽然该模型简单地分为阻力和推力两部分,但仍能得到令人满意的仿真结果。模拟速度与实验速度的变化趋势一致。在相同的频率下,所有的模拟速度都略大于实验速度,误差不超过20%。这种偏差主要是由于机器人固有的机械装配和水环境的不确定性造成的。此外,为了简化计算而忽略的游动速度也会通过影响尾鳍的有效攻角而对推力做出一定的贡献。
图8(a)情况1不同频率的模拟游泳速度;(b)前向游泳的模拟和实验游泳速度的比较
5 结论
综上所述,我们提出了一个简单而有效的基于CPG运动控制的水下仿生机器人推力产生与尾鳍振荡的关系模型。为了简化,将作用在机器人上的力分为推力和阻力。通过DPIV实验和力传感器实验确定了平均推力系数和瞬时推力系数。通过数值模拟和物理实验验证了所提出的动态模型的有效性。结果表明,特征速度是计算推力的有效因素。这个简单的模型将有助于设计仿生机器人。在未来的工作中,我们将更多地研究机器人尾幅、尾频与尾涡的双向相互作用。此外,还将建立一种运动优化方法,以提高机器人在水中的游泳能力。